Радіонуклідне забруднення макроміцетів

Результати досліджень, проведених у національному парку Евре Дівідал (0vre Dividalen) на півночі Норвегії, свідчать, що дикорослі їстівні гриби та ягоди мали близькі рівні акумульованого 210Pb, але рівні накопичення 210Po та 40K у грибах були вищими, ніж у ягодах. Середні концентрації активності 210Po в L. scabrum досягали 94, а в L. versipelle — 198 Бк/ кг с.м., що набагато перевищувало… Читати ще >
Радіонуклідне забруднення макроміцетів (реферат, курсова, диплом, контрольна)
Радіонуклідне забруднення макроміцетів
Планетарні масштаби ядерних аварій та їх тяжкі екологічні наслідки зумовлюють необхідність довготривалого моніторингу довкілля. Ключова роль мікобіоти полягає не лише в її участі в процесах біогенної міграції радіонуклідів у ґрунтах, а й у іммобілізації та утримуванні значної кількості радіонуклідів ґрунтовою міцеліальною біомасою [1−4].
Численні публікації останніх десятиліть присвячені високим акумулятивним властивостям шапинкових грибів щодо важких металів, радіонуклідів природного і техногенного походження [5−43]. Уперше явище накопичення макроміцетами радіонуклідів внаслідок їх глобальних викидів після випробувань ядерної зброї було встановлено Г. Грютером у 1963 р. У зразках шапинкових грибів із Західної Німеччини було виявлено вибіркове накопичення не лише 137Cs, а й інших продуктів розпаду — 144Ce, 106Ru,90Rh і 90Sr [44]. У дочорнобильський період найвищу активність 137Cs (до 25,2 кБк/кг сухої маси) було зафіксовано австрійським ученим К. Хазельвандтером у Cortinarius armillatus [45]. Дослідження біоакумуляції та переносу радіонуклідів з ґрунту у плодові тіла макроміцетів після чорнобильської катастрофи були зосереджені переважно на радіоцезії (137Cs, у перші роки після аварії у поєднанні з 134Cs) і частково на 90Sr, оскільки їх було визнано головними дозоутворювальними радіонуклідами. Цим пояснюється недостатня вивченість акумуляції макроміцетами інших радіонуклідів як природного, так і техногенного походження, інтерес до яких посилився лише в останні роки.
Серед радіонуклідів природного походження у грибах спостерігаються найвищі рівні 40К, який у плодових тілах розподіляється нерівномірно, кількісно знижуючись у послідовності шапинка > ніжка > пластинки або трубочки гіменофора > спори. Активність 40К, визначена у багатьох видів дикорослих макроміцетів Центральної Європи в період між 1984 і 1992 р., перебувала в межах 800−1500 Бк/кг сухої маси [30, 31]; у зразках грибів Українського Полісся, взятих у період 1990;2010 рр., — була на такому самому рівні, лише у деяких видів відзначено підвищені рівні цього ізотопа, зокрема у Boletus chrysenteron (українські назви грибів наведено в додатку) — до 11 000, у видів роду Amanita — до 7000, тоді як рівень 40К у ґрунтах з місцезростань грибів становив 100−700 Бк/ кг с.м. (рис. 1) [4, 18].
радіонуклід макроміцет забруднення.
![Активність K у плодових тілах макроміцетів у 1991 р. (зразки з Київської, Чернігівської та Житомирської областей) [4].](/img/s/9/58/2239858_1.jpg)
Рис. 1. Активність 40K у плодових тілах макроміцетів у 1991 р. (зразки з Київської, Чернігівської та Житомирської областей) [4].
Найвищий рівень активності 40К виявлено у Laccaria laccata — до 12 000 Бк/кг с.м. [46]. Мінімальні коефіцієнти накопичення (Кн дорівнює співвідношенню активності радіонукліда в плодовому тілі гриба до його активності у ґрунті/субстраті в точці збору) 40К (близько 2,0) були встановлені нами у сапротрофних і лігнотрофних видів — Macrolepiota procera, M. rachodes, Tricholomopsis rutilans, максимальні — у B. chrysenteron (98), Amanita citrina (63), Coltriciaperennis (48), Agaricus sylvaticus та A. rubescens (20).
У мінеральному складі плодових тіл шапинкових грибів калій є макроелементом, тому поглинання його з ґрунтів/субстратів (у тому числі 40К) неминуче. Зважаючи на те, що 40К, на відміну від 137Cs з радіоактивних опадів, рівномірно розподілений по вертикальному профілю лісових ґрунтів, припускають, що на його акумуляцію плодовими тілами рівень залягання міцелію не впливає [47].
Активність 210Pb (продукт розпаду природних ізотопів 238U, 226Ra, 222Rn) у грибах зазвичай на два порядки нижча, ніж 40К [31]. Максимальну активність 210Pb було виявлено в шапинках та цілих плодових тілах Boletus edulis, відповідно 58,9 і 36,6 Бк/кг с.м. [48]. Також у грибах з Польщі було знайдено природний ізотоп 210Po (продукт розпаду 238U) [33]. Слід зазначити, що у досліджених зразках 20 їстівних та неїстівних видів грибів максимальні рівні активності 210Po виявлено у B. edulis, Leccinum scabrum — до 40 Бк, а мінімальні 2,1−4,3 Бк/кг с.м. у Xerocomus badius (Boletus badius) і X. subtomentosus (B. subtomentosus) (до речі, обидва види є типовими акумуляторами радіоцезію). У шапинках рівні цього радіонукліда вищі, ніж у ніжках. За умови річного споживання людиною 5 кг свіжих плодових тіл B. edulis, отримана ефективна доза від 210Po досягає 37 мкЗв [33]. Автори зазначають, що активність 210Po в грибах була істотно вищою, ніж у продуктах харчування рослинного і тваринного походження. У роботі [49] також встановлено найвищі рівні 210Po у болетальних грибах (а саме, у Leccinum vulpinum), а низькі — у представників родини Russulaceae. При цьому розрахована річна доза від 210Po внаслідок споживання грибів в умовах Фінляндії становить близько 4 мкЗв.
Результати досліджень, проведених у національному парку Евре Дівідал (0vre Dividalen) на півночі Норвегії, свідчать, що дикорослі їстівні гриби та ягоди мали близькі рівні акумульованого 210Pb, але рівні накопичення 210Po та 40K у грибах були вищими, ніж у ягодах. Середні концентрації активності 210Po в L. scabrum досягали 94, а в L. versipelle — 198 Бк/ кг с.м., що набагато перевищувало активність цього радіонукліда у плодових тілах інших досліджених мікосимбіотрофних видів — Russula paludosa (4,7) і R. decolorans (7,4). З огляду на те, що болетальні види є визнаними акумуляторами селену (аналога полонію), автори роботи [50] висловили припущення, що значне перевищення рівнів акумуляції 210Po у видів роду Leccinum порівняно з Russula spp. пов’язане саме з цим фактом. Співвідношення активностей 210Po/210Pb в усіх досліджених зразках грибів завжди було більшим за одиницю. При цьому автори підкреслюють, що в цьому регіоні внесок 137Cs чорнобильського походження не такий вагомий, як у центральній частині Норвегії, тому доза (за умови високого рівня споживання лісової продукції), зумовлена переважно комбінацією 210Po, 210Pb і 40K, досягає 0,05 мЗв/рік за рахунок ягід і 0,50 мЗв/рік за рахунок грибів [50].
Нечисленні літературні дані свідчать про досить низький рівень активності 226Ra в плодових тілах макроміцетів з європейських країн [51, 52]. В Україні 226Ra і 228Ra ми виявили в 1991 р. у деяких зразках грибів, зокрема 226Ra у Pholiota destruens — 22 Бк/кг с.м. (м. Київ), 226Ra і 228Ra у Marasmius oreades — відповідно 22 і 15 Бк/кг с.м. (Київська обл., Обухівський р-н) і 228Ra у A. campestris — 71 Бк/кг с.м. (Чернігівська обл., Бобровицький р-н) [18].
Здатність грибів акумулювати природні ізотопи 234,238U і 228,230,232Th привертає особливу увагу. Дослідження, проведені у двох географічно віддалених (понад 250 км), але екологічно подібних екосистемах Іспанії з високою продуктивністю макроміцетів, показали, що TF (transfer factor, або коефіцієнт переносу) урану був у межах 0,043−0,049, а TF торію — 0,0300,62. Найвищі коефіцієнти біодоступності (визначаються так само, як і Кн) 234U і 238U спостерігалися у Hebeloma cylindrosporum (8,2 і 6,3), Amanita muscaria (6,0 і 4,5), Tricholoma terreum (4,0 і 3,8) і T. pessandatum (4,0 і 3,1).
Водночас найвищі коефіцієнти біодоступності 228,230,232Th встановлено для T. terreum (відповідно 64, 316 і 272), H. cylindrosporum (62, 371 і 360) та A. muscaria (38, 195 і 188). Порівняння коефіцієнтів переносу досліджених природних радіонуклідів з техногенними показало, що біодоступність торію близька до біодоступності цезію, а урану — вища, ніж у 90Sr, 239+240Pu і 241Am.
Отже, незважаючи на загальну достатньо низьку доступність торію і урану у ґрунтах, макроміцети все ж здатні до переносу і накопичення цих елементів. Автори роботи [53], враховуючи ці результати й отримані раніше дані щодо високих рівнів акумуляції радіоцезію Н. cylindrosporum та іншими представниками роду Hebeloma, вважають, що вони є надійними біоіндикаторами радіонуклідного забруднення довкілля.
Радіоізотоп 7Be, що генерується в атмосфері внаслідок її взаємодії з космічним випромінюванням високої енергії, до останнього часу було знайдено лише в деяких грибних зразках з Іспанії та Фінляндії [34, 54, 55]. Його надходження у плодові тіла пояснюють атмосферними опадами, при цьому рівень акумуляції визначається типом живлення макроміцетів, а саме — сапротрофні види накопичують високі й середні рівні 7Be, що пов’язано з розташуванням їх міцелію в багатому на гумус підстилковому шарі ґрунту [34]. Однак за даними T. Lonnroth зі співавторами, представники іншої екологічної групи — мікоризоутворювачі (Hygrophorus hypothejus, Russula paludosa, Hydnum rufescens, Lactarius rufus) є акумуляторами 7Be [54].
Добре відомо, що аварія 1986 р. на Чорнобильській АЕС призвела до значного поширення ряду радіоактивних елементів у Північній півкулі. Активна емісія радіонуклідів зі зруйнованого реактора у поєднанні зі змінюваними метеорологічними умовами і різноманіттям ландшафтів зумовили надзвичайно неоднорідний склад радіоактивних опадів на території України та суміжних держав.
кБк/кг с. м. 140.
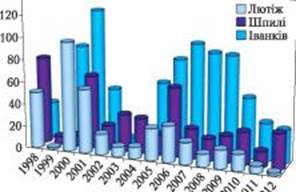
Рис. 2. Активність 137Cs у плодових тілах Boletus badius з місцезростань Київської області (м. Іванків, с. Шпилі Іванківського р-ну, с. Лютіж Вишгородського р-ну) Унаслідок аварії зафіксовано багаторазове збільшення вмісту радіонуклідів у дикорослих макроміцетах [4, 10, 11, 14, 16−18, 20, 23, 26, 30, 31, 56−60]. Автори повідомляли, що рівень природного ізотопа 40К у плодових тілах залишався достатньо стабільним (700−1000 Бк/кг с.м.), тоді як рівень 137Cs+134Cs зазвичай був вищим і значно варіював [9, 61, 62]. У перший післяаварійний період саме за присутністю 134Cs у поєднанні з 137Cs (у співвідношенні 1:2) можна було робити висновок про чорнобильське походження радіоцезію в грибах, оскільки активність 137Cs в плодових тілах макроміцетів також може бути пов’язана з глобальними випадіннями після випробувань ядерної зброї у 50−60-х роках минулого століття [7, 12, 41, 63].
Автори дослідження [1] показали, що понад 40% акумульованого радіоцезію зв’язано з гіфами, що переконливо свідчить про те, що у ґрунтах грибна біомаса може іммобілізувати значну кількість радіоцезію на невизначено довгий період. За даними, наведеними в [64], міцелій у верхньому органічному шарі ґрунту може містити до 50% 137Cs, локалізованого у 0−10-сантиметровому шарі лісових ґрунтів Швеції та України. Встановлено, що гриби здатні не лише безпосередньо зв’язувати або фіксувати радіонукліди, а й опосередковано впливати на форми, в яких вони знаходяться і переміщуються в лісових ґрунтах [3]. M. Steiner зі співавторами вважають, що гриби відіграють ключову роль у мобілізації, поглинанні та переносі поживних речовин і вочевидь є важливим фактором довгострокового утримування радіоцезію в органічному шарі ґрунту [2]. P. Kalac [31] дотримується подібних поглядів: гриби (як міцеліальні, так і ті, що утворюють плодові тіла) дуже ефективні в абсорбції радіонуклідів і є надзвичайно важливим компонентом у довгостроковій акумуляції завдяки довгоживучій і величезній мережі гіфів та грибної біомаси у верхніх шарах лісових ґрунтів. Дійсно, за оцінкою Л.Г. Бурової, біомаса макроміцетів з урахуванням міцелію у 5 разів перевищує надґрунтовий покрив [65]. Залежно від біологічних особливостей, зокрема глибини залягання (розташування) міцелію, макроміцети здатні «захоплювати» 137Cs як з поверхні ґрунту, підстилки, так і з більш глибоких шарів. Макроміцети, без сумніву, залучені до біогеохімічного круговороту 137Cs. У процесі розкладання плодових тіл, як і у випадку інших мінеральних елементів, гриби здатні рециркулювати радіоцезій з глибоких шарів ґрунту на поверхню лісової підстилки [40].
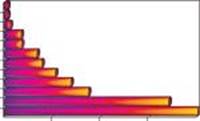
0 5000 10 000 15 000 20 000.
кБк/кг c. m.
Рис. 3. Активність 137Cs у плодових тілах макроміцетів з Чорнобильської зони (Старі Шепеличі, лісни-цтво, 2004.
Рівень накопичення радіонуклідів макроміцетами визначається комплексом чинників, серед яких, безумовно, найважливішим є кількісний та якісний склад аварійних викидів, рівень забруднення ґрунтів (субстратів) з місцезростань. Важливим також є фізико-хімічний склад та структура забруднених ґрунтів. G. Heinrich вважає, що накопичення радіоцезію макроміцетами залежить переважно від вмісту глини, мулу, піску, від обмінних катіонів та розподілу часточок різного розміру [66].
Є дані про вплив типу лісу на рівень акумуляції радіоцезію, зокрема, одні й ті самі види грибів у соснових лісах були більш забрудненими, ніж у мішаних [67]. Встановлено позитивну кореляцію між ступенем гідроморфності ґрунтів і вмістом радіонуклідів у грибах. Види, що зростають на гідроморфних лісових ґрунтах акумулятивних ландшафтів, накопичують 137Cs на порядок більше, ніж ті самі види, що зростають на автоморфних ґрунтах елювіальних ландшафтів [32, 60].
Вирішальну роль у рівнях біоакумуляції радіонуклідів відіграє глибина розташування міцелію окремих видів у ґрунті. Види, міцелій яких знаходиться у верхньому шарі ґрунту (зокрема, представники родів Collybia і Clitocybe), були найбільш забрудненими у перші роки після аварії, пік забруднення видів з глибоким заляганням міцелію (B. edulis) спостерігався лише через кілька років [15, 46, 59, 68−70].
Визначення коефіцієнтів накопичення (Кн) з ґрунтів у плодові тіла показало, що вони можуть досягати 4 порядків величин [71]. У наших дослідженнях максимальні Кн були зафіксовані у Paxillus sp. — до 1180, Lactarius helvus — до 855, Suillus luteus —766 та H. crustuliniforme — 514 (Київська обл., 1996, 1998 р., середній рівень поверхневого забруднення ґрунтів 137Cs — 37−185 кБк/м2). У посушливих умовах, за малої кількості опадів спостерігається зниження Кн радіоцезію. Динаміка акумулятивної здатності макроміцетів у часі подібна до затухаючого коливального процесу з піками у вологі роки і спадами у посушливі (рис. 2). І навпаки, підвищені Кн — до 500 (за відносно низької активності радіоцезію в ґрунтах) спостерігалися, зокрема, у макроміцетах, що зростали на заболочених, торф’янистих, низькогумусових ґрунтах (Волинська обл., Черемський заповідник) [38].
Надзвичайно високі рівні (до мільйонів Бк/ кг с.м.) акумуляції радіоцезію у плодових тілах деяких мікосимбіотрофних видів спостерігалися в зоні відчуження ЧАЕС: Gomphidius glutinosus — 3,78 МБк/кг с.м. (Старі Шепеличі, лісництво, 1992); Lactarius turpis — 13 і B. edulis — 1,56 МБк/кг с.м. (Новошепеличі, 1993); Paxillus involutus — до 15,71, Lactarius rufus — 12,58, Cantharellus cibarius — 12,13, Boletus badius —11,83, B. edulis — 5,11 МБк/ кг с.м. (Прип'ять, «Рудий ліс», 1996); Pluteus cervinus — 34,59, Leccinum scabrum — 5,46 Hvgrophoropsis auranliaea A nnillariclla inellea Amanita a Irina Tricholoma sp. Boletus badius ('antliarcllus cibarius Laclarius vclicrcus Stropharia aeruginosa Laclarius dcliciosus Laclarius turpis Paxillus involutus Boletus subtomentosus МБк/кг с.м. (Новошепеличі, 1996); Boletus subtomentosus — 20, P. involutus — 17 МБк/ кг с.м. (Старі Шепеличі, лісництво, 2004); L. turpis — 18,71 i Suillus luteus — 14,88 МБк/кг с.м. (Копачі)(рис. 3). Разом з тим Кн у макроміцетів, що зростають у таких сильно забруднених локалітетах, перебували в межах 1−50 [4, 26, 38, 72].
До останніх років, за нашими даними, активність радіоцезію у близько 60% плодових тіл цінних їстівних видів грибів-мікосимбіотрофів (Boletus spp., Suillus spp., Leccinum scabrum, Cantharellus cibarius, Lactarius spp., Tricholoma flavovirens та ін.), зібраних на території Українського Полісся (за рівнів забруднення >37 кБк/м2 [73]), перевищують гранично допустимі рівні, прийняті в Україні (2500 Бк/кг с.м.) (рис. 4) [4, 74]. Проте слід зазначити, що досить низькі рівні активності 137Cs спостерігаються в плодових тілах Agaricus spp., Macrolepiota procera, Armillariella mellea, Kuehneromyces mutabilis, Fistulina hepatica, Sparassis crispa, Trametes versicolor, Pleurotus ostreatus, Piptoporus betulinus, Lycoperdon spp., Calvatia utriformis, Langermannia gigantea. Остання група грибів, яка є достатньо безпечною з точки зору вмісту радіоцезію, на забруднених територіях потребує додаткового контролю щодо вмісту радіостронцію, з огляду на те, що серед них багато лігнотрофів, яким властиве підвищене накопичення радіостронцію з рослинних субстратів [4].
За даними дослідження [67], активність у 2030% грибів та ягід у Гомельській області вища за допустимий рівень, який у Білорусі становить 370 і 185 Бк/кг сирої ваги відповідно.
Piploporus belli linns Нуріюіота fasciculate Amanita panllicrina СШосуЬе iiifunilibuliforiiiix Лиши На muscaria Boletus edulis Macrolepiota rliacodes Anuillariella mellea Amanita citrina Triclioloinopsis rutilaus Suillus gran и lams Macrolepiota procera Tricliolonia fluvobriinneuni Tricholoma Jlavovirens Amanita rubes cats Russula xerainpelina Cantliarellns cibarius Lada rius voleinus Suillus luteus Boletus badius I. a eta rius nirpis Paxillus involutns Russula vesca Russula cyanoxantha La da rius liehns Lepista gib’at «or fin a rius praestans

0 50 100 200 250.
кБк/кг c. m.
Рис. 4. Активність радіоцезію в плодових тілах макроміцетів у 2010 р. (с. Карпилівка, Чернігівська обл., Козелецький р-н, рівень поверхневого забруднення ґрунтів 137Cs >37 Бк/кг с.м.).
На акумуляцію радіоцезію макроміцетами істотно впливає екологічна приуроченість видів, а саме — стратегія живлення: зменшення спостерігається у послідовності від мікосимбіотрофних сапротрофних лігнотрофних видів [4, 23, 26, 38, 70]. До останнього часу такої специфічної тенденції щодо акумуляції радіостронцію не виявлено.
Зазвичай рівні радіостронцію в плодових тілах макроміцетів у середньому в сотні разів менші, ніж радіоцезію. За нашими даними, у 2004 р. в зразках грибів із 30-кілометрової зони ЧАЕС співвідношення 137Cs/90Sr було в межах 10— 104, при середньому значенні 103. У мікосимбіотрофного виду Cantharellus cibarius з «Рудого лісу» активність 90Sr досягала 10 240 Бк/кг с.м. (1996 р.). Найвищу активність радіостронцію було виявлено у сапротрофа Stropharia aeruginosa — 13 000 та мікосимбіотрофа Lactarius turpis — 11 000 Бк/кг с. м (рис. 5). Слід зазначити, що допустимий рівень 90Sr для сухих грибів в Україні становить 250 Бк/кг с.м. [74].
Як живі, так і мертві гіфи здатні до швидкого, не залежного від метаболізму, зв’язування радіонуклідів клітинною стінкою, екстрацелюлярними полісахаридами або іншими компонентами з біосорбційними властивостями [12]. Автори зазначають, що суттєві відмінності у будові клітинної стінки різних видів зумовлюють значну варіабельність в акумулювальній здатності грибів. Відомо, що більшість макроміцетів мають обмежену здатність до накопичення стабільного цезію. Методом атомно-абсорбційної спектроскопії було досліджено його вміст у плодових тілах 433 видів дикорослих грибів Європи і показано, що середній вміст цього елемента становив 7 мг/ кг с.м., при цьому надзвичайно високі концентрації спостерігалися у представників родини Cortinariaceae (особливо у Cortinarius alboviolaceus), високі — у Clavariaceae, Rhodophyllaceae, Strophariaceae, а у представників Helvellaceae та Lycoperdaceae — низькі [75].
Отже, можна припустити, що процеси накопичення радіонуклідів макроміцетами повинні відбуватися певною мірою аналогічно накопиченню стабільних елементів. Справді, L.R. Bakken і R.A. Olsen [13], вивчаючи накопичення радіоцезію макроміцетами Норвегії, встановили позитивну кореляцію між концентраціями радіоцезію та нерадіоактивного цезію і негативну — з рівнем рН ґрунту. Перевірка авторами гіпотези щодо механізму надходження цезію у плодові тіла, яка основана на передбачуваній здатності транспортних ензимів клітинних мембран розпізнавати іони Cs+ і K+, показала відсутність цього розпізнавання. Міжвидові відмінності у поглинанні радіоцезію та стабільного цезію автори пояснюють насамперед просторовим розміщенням цих елементів у ґрунтових шарах та рівнем спорідненості до Cs. Важливим, на нашу думку, є припущення, що гриби, змінюючи рН середовища, здатні вивільняти радіоцезій, який до цього перебував у зв’язаному стані.
Разом з тим дослідження впливу стабільного цезію, що знаходиться у лісовій підстилці, на рівень переносу радіоцезію у плодові тіла макроміцетів спростували наявність такої кореляції у зразках з місцезростань Польщі з різним ступенем промислового забруднення [76].
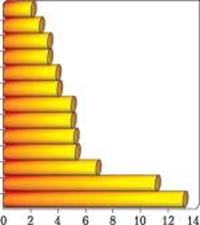
Рис. 5. Активність 90Sr у плодових тілах макроміцетів з Чорнобильської зони (Старі Шепеличі, лісництво, 2004) кБк/кг с. м.
Культуральні експерименти з Pleurotus ostreatus [77] показали, що 137Cs і стабільний Cs активно поглиналися грибом залежно від концентрації 137Cs або Cs у живильному середовищі. Автори встановили, що накопичення 137Cs плодовими тілами пригнічувалося К та/або Cs, що підтверджує здатність грибів до поглинання 137Cs через К-транспортні системи.
Рівні акумуляції стабільного цезію грибами подібні до тих, що спостерігаються у судинних рослин. Проте у випадку радіоцезію спостережувані величини як мінімум на порядок вищі [30]. Різна поведінка природного і радіоактивного цезію у цьому випадку пов’язана з їх нерівноважним станом в екосистемі та різним ступенем їх доступності [30, 78].
Результати експериментальних досліджень поведінки ізотопів цезію (137Cs і 133Cs) та їх «двійників» — калію та рубідію в системі переносу ґрунт-гриби-рослини, проведених у лісових екосистемах Швеції, показали, що гриби накопичують значно більше K та Ru, ніж стабільного 133Cs, причому у плодових тілах концентрація цих елементів у середньому була на порядок вищою, ніж у міцелії [79]. Спорідненість до цих елементів у грибах (плодових тілах і міцелії) знижувалася у ряду Rb+ > K+ > > Cs+ з відносним співвідношенням 100:57:32, що було близьким до більш ранніх даних японських дослідників — 100:88:50 [25].
Хоча широкомасштабна ремедіація територій, забруднених радіонуклідами і особливо 137Cs, є проблематичною, K. Rosen зі співавторами показали, що одноразова обробка лісового ґрунту калійним добривом (KCl) у 1992 р. довгостроково (навіть через 17 років після застосування) впливала на біоакумуляцію 137Cs видами Cortinarius semisanguineus, Lactarius ru-
Boletus versicolor Lactarius vellereus Lactarius deliciosus Boletus badius CKtocybe infundibuliformis Amanita citrina Hygrophoropsis aurantiaca Canlharellus abortus Ралі Hus involutus Russula xerampelina ArmiUariella mellea Lactarius turpis Stropharia aeruginosa fus, C. caperatus і Suillus variegatus, радіоактивність у яких зменшувалася від 21 до 58% [80].
G. Heinrich описав нерівномірність розподілу 137Cs у плодових тілах грибів у вигляді такої послідовності — пластинки > м’якуш шапинки > ніжка [81]. Дослідження біоакумуляції ряду природних та антропогенних радіонуклідів у серії культуральних експериментів з Pleurotus eryngii, вирощеними у контрольованих лабораторних умовах, та з Tricholoma equestre, зібраними у напівприродній екосистемі, показали, що ступінь зрілості відіграє важливу роль у поглинанні та розподілі радіонуклідів у плодових тілах цих видів [35]. Максимальна інкорпорація 134Cs і 85Sr спостерігається у зрілих плодових тілах, потім знижується з їх старінням. У процесі дозрівання загальна активність 85Sr, 134Cs i 60Co зростає у шапинці та пластинках з відповідним зниженням у ніжці гриба. Для обох досліджених видів було показано, що радіоцезій і калій переважно локалізуються у шапинці з пластинками, а 226Ra — у пластинках.
Оскільки у досліджених макроміцетів розподіл 239,240Pu, 234,238u i 23o,232Th різнився, автори припустили, що він є видоспецифічним [35].
Численні дані щодо вмісту радіоцезію в грибах Європи, незважаючи на деякі протиріччя, свідчать про видоспецифічність накопичення. Найвищі рівні активності та коефіцієнтів переносу (TF) виявлено у мікосимбіотрофних видів, зокрема у представників родин Cortinariaceae і Boletaceae [11, 12, 14, 20, 21, 23, 26−28, 48, 61, 82].
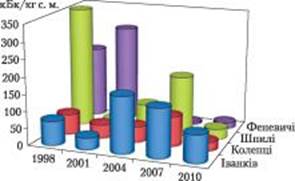
Рис. 6. Динаміка активності 137Cs у плодових тілах біоіндикатора Paxillus involutus з міцезростань Київської області: південні околиці м. Іванків, околиці с. Коленці, Шпилі та Феневичі (Іванківський р-н).
K. Haselwandter першим висловив припущення, що гриби акумулюють 137Cs видоспецифічно [83]. Незважаючи на значну варіабельність даних, саме ця особливість макроміцетів може бути використана для моніторингу радіоактивності довкілля [6, 12, 38, 55]. Спостережуване в деяких мікосимбіотрофних макроміцетах суттєве перевищення рівнів забруднення порівняно з лісовою підстилкою (основним депо радіонуклідів) та іншими компонентами екосистем, надає їм особливого статусу в системі біоіндикації. Серед видівгіперакумуляторів радіоцезію, надійними інформативними індикаторами впродовж усього післячорнобильського періоду залишаються рекомендовані раніше поширені в лісових екосистемах України Boletus badius, Lactarius rufus i Paxillus involutus (рис. 6) [4, 28, 38].
Міжвидові відмінності у накопиченні 137Cs у зразках макроміцетів, зібраних з тих самих місцезнаходжень, можуть досягати двох порядків величин. Цікавим є приклад видів роду Leccinum, зібраних у локалітетах з різним рівнем радіонуклідного забруднення. Активність радіоцезію у плодових тілах Leccinum scabrum завжди в рази вища, ніж у L. aurantiacum [4]. Видоспецифічність накопичення може бути пов’язана передусім з хімічними властивостями деяких компонентів плодових тіл. Так, підвищені рівні 137Cs, які зазвичай спостерігаються у B. badius, деякі автори пояснюють утворенням комплексу цього радіонукліда з двома коричневими пігментами кутикули шапинок — бадіоном, А та норбадіоном A [84, 85]. D. Aumann зі співавторами стверджують, що подібний механізм відповідає за акумуляцію радіоцезію також у Boletus erythropus і B. mirabilis [86]. Результати наших досліджень свідчать, що B. edulis з Народицького р-ну (околиці відселеного с. Христинівка) у плодових тілах з темнозабарвленими шапинками активність 137Cs становила 95 800 Бк/кг с.м., що майже у 3,25 раза вище, ніж у плодових тіл зі світлим кольором шапинок (29 400 Бк/кг с.м.). Це, вочевидь, свідчить про важливу роль пігментної складової у процесах біоакумуляції радіонуклідів.
Крім радіоцезію та радіостронцію в плодових тілах макроміцетів з 30-кілометрової зони ЧАЕС у 1995 р. ми виявили також 144Ce — 1071 (Leccinum scabrum, с. Стара Красниця), 144Ce і 154Eu — відповідно 310,1 і 31,3 кБк/кг с.м. (Lactarius turpis, c. Новошепеличі) [4, 72].
Випробування ядерної зброї та чорнобильска катастрофа є основними джерелами високотоксичного 241Am у довкіллі. За даними, наведеними в роботі [87], активність 241Am у грибах із забруднених регіонів Фінляндії в середньому становить 7,5 мБк/кг, проте коефіцієнт переносу цього радіонукліда значно вищий, ніж Pu, що свідчить про його вищу біодоступність. Плутоній у макроміцетах України було виявлено в межах 2,96−82,8 Бк/кг с.м., причому найвищі його концентрації спостерігалися в Paxillus involutus і Cantharellus cibarius. В інших країнах (Фінляндія, Японія, Польща, Іспанія) активність цього радіонукліда була значно нижчою (0,0009−0,164 Бк/кг с.м.), а найвищі рівні 241Am виявлено у плодових тілах Clitocybe sp., Hebeloma cylindrosporum і Lycoperdon perlatum [41, 55, 87−90]. Слід зазначити, що вперший післяаварійний період у макроміцетах було зафіксовано й інші короткоіснуючі радіонукліди, такі як 110Ag, 125Sb, 103Ru і 131I [55].
Цікавий методичний підхід для вирішення проблем очищення технологічних та природних вод від радіонуклідів запропонували Д.В. Дементьєв зі співавторами [91], продемонструвавши, що біомаса живого мицелію Pleurotus ostreatus здатна ефективно (>70%) сорбувати розчинений 241Am. Більша частина (90%) накопиченого в міцелії 241Am зв’язувалася зі структурними полісахаридами клітинних стінок. Показано, що питома активність радіонукліда в полісахаридах клітинних стінок була у 3,5 раза вищою, ніж у вихідній біомасі міцелію P. ostreatus.
До останнього часу у відкритому доступі є лише нечисленні відомості щодо біосорбції радіонуклідів макроміцетами внаслідок ядерної аварії на Фукусімі. Після цієї події (березень 2011 р.) до кінця 2014 р. з метою мінімізації дози внутрішнього опромінення було проведено національний широкомасштабний моніторинг харчових продуктів з різних префектур Японії, який підтвердив перевищення допустимих рівнів 137Cs, передусім у грибах (максимальна виявлена активність сягала 28 000 Бк/кг) і сушених овочах. Повідомляється, що пік забруднення у сушених шіїтаке (Lentinus edodes) спостерігали з вересня 2011 до кінця березня 2012 р. [92]. K. Nakashima зі співавторами показали, що у 36,4% дикорослих грибів, зібраних в околицях с. Каваучі (30 км від станції), активність становила близько 1000 Бк/кг, а максимальну активність 134Cs 5432,7 і 137Cs 11 616,2 Бк/кг с.м. було спостережено у Cortinarius salor Fr. При цьому ефективна доза для дорослих, розрахована на середньорічне споживання грибів, була в межах 0,11−1,6 мЗв [93].
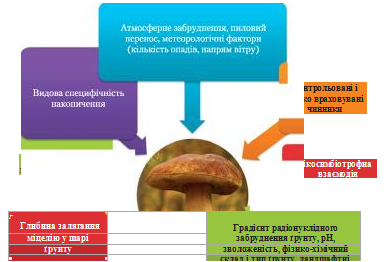
Аналіз літературних і авторських даних дозволив згрупувати абіотичні та біотичні чинники, що визначають ступінь радіаційного забруднення макроміцетів (рис. 7).
Загалом рівень біоакумуляції радіонуклідів макроміцетами залежить від конкретної радіоекологічної ситуації в місці збору, видоспецифічності та екологічної приуроченості виду гриба. Разом з тим, значний рівень варіабельності, спостережуваний в отриманих даних, свідчить про комплексний результуючий вплив певних (враховуваних і неконтрольованих факторів), що ускладнює прогнозну оцінку рівнів забруднення.
Незважаючи на те, що з моменту аварії на Чорнобильській АЕС минуло вже понад 30 років, й дотепер зберігається небезпека для населення, пов’язана з внутрішнім опроміненням унаслідок вживання продуктів, забруднених радіонуклідами, зокрема дикорослих грибів та ягід. Найцінніші їстівні види макроміцетів з порядку Boletales є акумуляторами не лише радіоцезію, а й природних ізотопів — 210Pb, 210Po, 40K. У країнах Європи щорічне споживання дикорослих грибів може сягати кількох кілограмів на душу населення [31], тоді як для жителів України (особливо Українського Полісся) лісові гриби не лише становлять значну частину традиційного раціону, а й забезпечують додатковий заробіток сільському населенню.
Єдине розумне вирішення проблеми — підвищення обсягів промислового виробництва цінних їстівних та лікарських видів грибів на попередньо перевірених субстратах. Проте в останні роки значне зниження рівня життя населення неминуче змушує наших співгромадян нехтувати цілком обґрунтованою небезпекою і збільшувати вживання саме «безкоштовних» лісових грибів.
Враховуючи наведені вище дані, очевидною стає необхідність довгострокового моніторингу з урахуванням вагомого внеску мікобіоти у процеси міграції та перерозподілу радіонуклідів для комплексної оцінки актуальної радіоекологічної ситуації і мінімізації негативних наслідків.
В основі сучасного підходу до оцінювання екологічного стану довкілля лежить антропоцентрична концепція. Якість життя та здоров’я людини в умовах зростаючого техногенного, у тому числі радіаційного, пресингу повинні стати пріоритетними напрямами наукових досліджень.
Література
- 1. Dighton J., Clint G.M., Poskitt J. Uptake and accumulation of 137Cs by upland grassland soil fungi: a potential pool of Cs immobilization. Mycol. Res. 1991. 95(9): 1052.
- 2. Steiner M., Linkov I., Yoshida S. The role of fungi in the transfer and cycling of radionuclides in forest ecosystems. J. Environ. Radioact. 2002. 58(2−3): 217.
- 3. Gadd G.M. Influence of microorganisms on the environmental fate of radionuclides. Endeavour. 1996. 20: 150.
- 4. Grodzinskaya A.A., Syrchin S.A., Wasser S.P., Kuchma N.D. Macromycetes accumulative activity in radionuclide contamination conditions of the Ukraine territory. In: Zhdanova N.N. (ed.). Mycobiota of Ukrainian Polesie: Consequences of the Chernobyl disaster. (Kyiv: Naukova Dumka, 2013). P. 217−260; 368−373.
[Гродзинская А.А., Сырчин С. А., Вассер С. П., Кучма Н. Д. Аккумулятивная активность макромицетов в условиях радионуклидного загрязнения Украины. В кн.: Микобиота Украинского Полесья: Последствия чернобыльской катастрофы (под общ. ред. Н.Н. Ждановой). К.: Наукова думка, 2013. C. 217−260; 368−373.].
- 5. Tyler G. Metals in sporophores of Basidiomycetes. Trans. Br. Mycol. Soc. 1980. 74: 41.
- 6. Haselwandter K., Berreck M., Brunner P. Fungi as bioindicators of radiocesium contamination: preand post-Chernobyl activities. Trans. Br. Mycol. Soc. 1988. 90: 171.
- 7. Haselwandter K., Leyval C., Sanders F.E. Impact of arbuscular mycorrhizal fungi upon plant uptake of heavy metals and radionuclides from soil. In: Gianianazzi S., Schuepp H. (eds.). Impact of Arbuscular Mycorrhizas on Sustainable Agriculture and Natural Ecosystems. (Basel: Birkhauser, 1994). P. 179−189.
- 8. Dietl G., Breitig D. Radioaktives Caesium in Pilzen aus dem Raume Schwabisch Gmund. Zeitschrift fur Mykologie. 1988. 54(1): 109.
- 9. Dighton J., Horrill A.D. Radiocaesium accumulation in the mycorrhizal fungi Lactarius rufus and Inocybe longicystis, in upland Britain, following the Chernobyl accident. Trans. Br. Mycol. Soc. 1988. 91(2): 335.
- 10. Teherani D.K. Determination of 134Cs and 137Cs radioisotopes in various mushrooms from Austria one year after the Chernobyl incident. J. Radioanal. Nucl. Chem. Lett. 1988. 6(6): 401.
- 11. Haselwandter K., Berreck M. Radiocesium accumulation in fruitbodies of Basidiomycetes collected in the province of Parma, Italy Fungi atque loci Natura, Atti del IV Convegno Intern. micol. del 27−30 IX 1987. Borgo Val di Taro, Italy, 1989. P 89−92.
- 12. Haselwandter K., Berreck M. Accumulation of radiocesium in fungi. In: Winkelmann G., Winge D.R. (eds.). Metal ions in fungi. (New York; Basel; Hong Kong: Marcel Dekker, 1994). P 259−277.
- 13. Bakken L.R., Olsen R.A. Accumulation of radiocaesium in fungi. Can. J. Microbiol. 1990. 36: 704.
- 14. Bem H., Lasota W., Kusmierek E., Witusik M. Accumulation of 137Cs by mushrooms from Rogozno area of Poland over the period 1984;1988. J. Radioanal. Nucl. Chem. Lett. 1990. 145(1): 39.
- 15. Fraiture A., Guillite O., Lambinon J. Interest of fungi as bioindicators of the contamination in forest ecosystems. In: Desmet G., Nassimbeni P, Belli M. (eds.). Transfer of radionuclides in natural and semi-natural environments. (London; New York: Elsevier Applied Science, 1990). P 477−484.
- 16. Wasser S. P, Grodzynska G.A., Lyugin V.O. Cs-134 and Cs-137 content in higher Basidiomycetes of Ukrainian Polissya. Ukrainian Botanical Journal. 1991. 48(5): 14.
[Вассер С.П., Гродзинська Г. А., Люгін В.О. Вміст Cs-134 і Cs-137 в вищих Basidiomycetes Українського Полісся. Укр. ботан. журн. 1991. T. 48, № 5. С. 14−19.].
17. Wasser S. P, Grodzynska G.A., Lyugin V.O. Uptake of radioactive elements by macromycetes of Ukrainian Polissya. Ukrainian Botanical Journal. 1992. 49(5): 79.
[Вассер С.П., Гродзинська Г. А., Люгін В. О. Накопичення радіоактивних елементів макроміцетами Українського Полісся. Укр. ботан. журн. 1992. T. 49, № 5. С. 79−86.].
- 18. Wasser S. P, Bolukh V.O., Brun' G.O., Virchenko V.M., Grodzynska G.A., Kondratyuk S.Ya., Navrotska I.L., Stupina V.V., Tsarenko PM. Uptake of radionuclides by cryptogame plants and higher fungi of Ukraine. (Kyiv, 1995). [Вассер С.П. та ін. Накопичення радіонуклідів споровими рослинами і вищими грибами України. Київ, 1995.]
- 19. Mietelski J.W., La Rosa J.L., Ghods A. 90Sr and 229+240pu, 238Pu, 241Am in some samples of mushrooms and forest soils from Poland.J. Radioanal. Nucl. Chem. 1993. 179: 243.
- 20. Mietelski J.W., Jasinska M., Kubicka B., Kozak K., Macharski P Radioactive contamination of Polish mushrooms. Sci. Total Environ. 1994. 157: 217.
- 21. Mietelski J.W., Macharski P, Jasinska M., Broda R. Radioactive contamination of forests in Poland. Biol. Trace Elem. Res. 1994. 43−45: 715.
- 22. Smith M.L., Taylor H.W., Sharma H.D. Comparison of the Post-Chernobyl 137Cs contamination of mushrooms from Eastern Europe, Sweden, and North America. Appl. Environ. Microbiol. 1993. 59(1): 134.
- 23. Wasser S. P, Grodzinskaya A.A. Content of Radionuclides in Macromycetes of the Ukraine in 1990;1991. In: Fungi of Europe: Investigation, Recording and Conservation. (Royal Botanic Gardens, Kew, 1993). P 189−210.
- 24. Yoshida S., Muramatsu Y. Concentrations of radiocesium and potassium in Japanese mushrooms. Environ. Sci. 1994. 7(1): 63.
- 25. Yoshida S., Muramatsu Y. Concentrations of Alkali and Alkaline Earth Elements in mushrooms and plants collected in a Japanese pine forest, and their relationship with 137Cs. J. Environ. Radioact. 1998. 41(2): 183.
- 26. Grodzinskaya A.A., Berreck M., Wasser S. P, Haselwandter K. Radiocesium in fungi: Accumulation pattern in the Kiev district of Ukraine including the Chernobyl zone. Sydowia. 1995. 10: 88.
- 27. Grodzinskaya A.A., Syrchin S.A., Kuchma N.D. Higher Basidiomycetes as bioindicators for radiocaesium contamination of territory of Ukraine. In: Botany and Mycology: modern horizons (to memory of Academician A.M. Grodzinsky (1926;1988). (Kyiv: Academperiodika, 2007). P 263−275.
- 28. Grodzinskaya A.A., Syrchin S.A., Kuchma N.D., Bilay V.T. Radioactive contamination of Ukrainian wild-growing mushrooms. In: Mushroom Biology and Mushroom Products: Proc. 7th Int. Conf. (ICMBMP7). Arcachon, 2011. P 566−572.
- 29. Fedorov V.N., Yeliashevich N.V. Accumulation of radionuclides in bodies of macromycetes. Radiation Biology. Radioecology. 2000. 40(6): 702.
[Федоров В.Н., Елиашевич Н. В. Аккумуляция радионуклидов в плодовых телах макромицетов. Радиац. биология. Радиоэкология. 2000. Т. 40, № 6. С. 702−709.].
- 30. KalaC P A review of edible mushroom radioactivity. Food Chem. 2001. 75: 29.
- 31. Kalac P Radioactivity of European wild growing edible mushrooms. In: Andres S., Baumann N. (eds). Mushrooms: Types, Properties and Nutrition. (New York, Nova Sci. Publ., 2012). P 215−230.
- 32. Scheglov A.I. Biogeochemistry of technogenous radionuclides in forest ecosystems. (Moscow: Nauka, 1999).
[Щеглов А. И. Биогеохимия техногенных радионуклидов в лесных экосистемах. М.: Наука, 1999.].
- 33. Skwarzec B., Jakusik A. 210Po bioaccumulation by mushrooms from Poland. J. Environ. Monit. 2003. 5: 791.
- 34. Baeza A., Hernandez S., Guillen F, Moreno G., Manjon J.L., Pascual R. Radiocaesium and natural gamma emitters in mushrooms collected in Spain. Sci. Total Environ. 2004. 318(1): 59.
- 35. Baeza A., Guillen F.J., Salas A., Manjon J.L. Distribution of radionuclides in different parts of a mushroom: Influence of the degree of maturity. Sci. Total Environ. 2006. 359(1): 255.
- 36. Ban-Nai T., Muramatsu Y., Yoshida S. Concentrations of 137Cs and 40K in mushrooms consumed in Japan and radiation dose as a result of their dietary intake. J. Radiat. Res. 2004. 45(2): 325.
- 37. Dvorak P, Kunova V., Benova K., Ohera M. Radiocesium in mushrooms from selected locations in the Czech Republic and the Slovak Republic. Radiat. Environ. Biophys. 2006. 45(2): 145.
- 38. Grodzynska A., Syrchyn S., Kuchma M., Konischuk V. Macromycetes — bioindicators of radiocaesium contamination of Ukrainian forest ecosystems. Visn. Nac. Akad. Nauk Ukr. 2008. (9): 26.
[Гродзинська Г. А., ^рчін С.О., Кучма М. Д., Коніщук В.В. Макроміцети-біоіндикатори забруднення радіоцезієм лісових екосистем України. Вісник НАН України. 2008. № 9. C. 26−37.].
- 39. Dementyev D.V., Bolsunovsky A.Ya. Accumulation of artificial radionuclides by edible wild mushrooms and berries in the forests of the central part of the Krasnoyarskii Krai. Radioprotection. 2009. 44(5): 115.
- 40. Falandysz J., Borovicka J. Macro and trace mineral constituents and radionuclides in mushrooms: health benefits and risks. Appl. Microbiol. Biotechnol. 2013. 97: 477.
- 41. Lehto J., Vaaramaa K., Leskinen A. 137Cs, 233,240Pu and 241Am in boreal forest soil and their transfer into wild mushrooms and berries. J. Environ. Radioact. 2013. 116: 124.
- 42. Falandysz J., Zalewska T., Apanel A., Drewnowska M., Kluza K. Evaluation of the activity concentrations of 137Cs and 40K in some Chanterelle mushrooms from Poland and China. Environ. Sci. Pollut. Res. 2016. 23(19): 20 039.
- 43. Falandysz J., Zhang J., Zalewska T. Radioactive artificial 137Cs and natural 40K activity in 21 edible mushrooms of the genus Boletus species from SW China. Environ. Sci. Pollut. Res. 2017. 24(9): 8189.
- 44. Grater H. Radioactive fission product 137Cs in mushrooms in W. Germany during 1963;1970. Health Physics. 1971. 20: 655.
- 45. Haselwandter K. Radioactives Casium (137Cs) in Fruchtkorpern verschiedener Basidiomycetes. Z. Pilzkunde. 1977. 43: 323.
- 46. Mietelski J.W., Dubchak S., Blazej S., Anielska T., Turnau K. 137Cs and 40K in fruiting bodies of different fungal species collected in a single forest in southern Poland. J. Environ. Radioact. 2010. 101(9): 706.
- 47. Krolak E., Kwapulinski J., Fischer A. 137Cs and 40K isotopes in forest and wasteland soils in a selected region of eastern Poland 20 years after the Chernobyl accident. Radiat. Environ. Biophys. 2010. 49: 229.
- 48. Malinovska E., Szefer P, Bojanowski R. Radionuclide content in Xerocomus badius and the other commercial mushrooms from several regions of Poland. Food Chem. 2006. 97(1): 19.
- 49. Vaaramaa K., Solatie D., Aro L. Distribution of 210Pb and 210Po concentrations in wild berries and mushrooms in boreal forest ecosystems. Sci. Total Environ. 2009. 408: 84.
- 50. Gwynn J. P, Nalbandyan A., Rudolfsen G. 210Po, 210Pb, 40K and 137Cs in edible wild berries and mushrooms and ingestion doses to man from high consumption rates of these wild foods. J. Environ. Radioact. 2013. 116: 34.