Роль системи глутамінової кислоти

Цей фермент менш активний в мозку, ніж в печінці. Він присутній у мітохондріях та потребує піридиннуклеотидних кофакторів і активується АДФ. Реакція зворотня, проте рівновага зміщена у бік прямої реакції, тобто синтезу глутамінової кислоти. Таким чином, у головному мозку глутаматдегідрогеназна реакція приймає участь не стільки в окисленні глутамата, скільки у синтезі його з б-кетоглутарату. Вона… Читати ще >
Роль системи глутамінової кислоти (реферат, курсова, диплом, контрольна)
В клітинах головного мозку має місце активний метаболізм амінокислот. Концентрація амінокислот в мозку майже в 8 разів вища, ніж у плазмі крові та у печінці. Особливо високим є рівень глутамата (5−10 мМ) і аспартата (1−3мМ). Ці амінокислоти утворюються в реакції трансамінування із проміжних метаболітів цитратного циклу, б-кетоглутарату і оксалоацетату. В тканинах мозку інтенсивно проходять метаболічні перетворення амінокислот — окислювальне дезамінування, трансамінування, перетворення радикалу. Особливо важливим для нормального функціонування головного мозку є реакція декарбоксилювання глутамату, в результаті якої утворюється г-аміномасляна кислота (ГАМК). Біосинтез і деградацію глутамату можно розглядати як шлях цитратного циклу (рис 3). ГАМК-шунт є характерним для клітин центральної нервової системи, але не відіграє суттєвої ролі в інших тканинах. Деякі амінокислоти, наприклад гліцин, аспартат, глутамат, ГАМК, таурин виконують функцію нейромедіаторів.
Вони зберігаються в синапсах і виділяються при надходженні нервового імпульса.
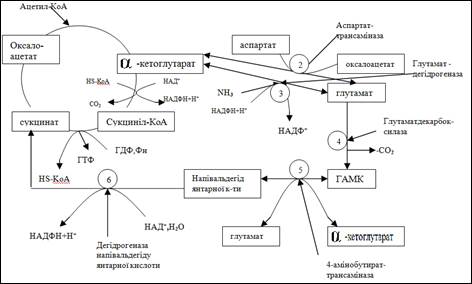
Рисунок 3. Метаболізм глутамата.
Глутамат, глутамін, ГАМК, аспартат, N-ацетиласпартат є домінуючими в кількісному відношенні, на їх долю приходиться 2/3 амінного азоту амінокислот головного мозку. В спинному мозку концентрація глутамата і споріднених амінокислот також, як і в головному, вище інших амінокислот. В периферийних нервах хребетних тварин вміст глутамата, глутаміна, N-ацетиласпартата значно менший чим в мозку, а ГАМК повністю відсутня. Особливістю метаболізму глутамата в нервовій тканині є її тісний зв`язок з інтенсивним функціонуванням в цьому органі цикла трикарбонових кислот (ЦТК). Це дозволяє вважати його проміжним продуктом енергетичного метаболізму. Основний енергетичний субстрат мозку — глюкоза — швидко перетворюється в амінокислоти. Так, вже через 30 хв. після ін'єкції міченої глюкози більше 70% приходиться на долю глутамата та його похідних. Цьому сприяють дуже швидке взаємоперетворення глутамата і б-кетоглутарата в ЦНС. Високий рівень інкорпорації радіоактивності із глюкози в амінокислоти мозку виявився підставою для ствердження, що утилізація глюкози в цьому органі, в значній мірі, протікає через біосинтез й окислення амінокислот. Безпосереднім попередником для синтезу глутамата в мозку є б-кетоглутарова кислота (рис. 4), яка здатна перетворюватися в глутамат шляхом прямого відновлювального амінування з участю глутаматдегідрогенази або шляхом трансамінування. Глутаматдегідрогеназа каталізує наступну реакцію:
бкетоглутарат + НАДН2 (НАДФН2) + NH3— глутамат + Н2О +.
+ НАД+(НАДФ+).
Цей фермент менш активний в мозку, ніж в печінці. Він присутній у мітохондріях та потребує піридиннуклеотидних кофакторів і активується АДФ. Реакція зворотня, проте рівновага зміщена у бік прямої реакції, тобто синтезу глутамінової кислоти. Таким чином, у головному мозку глутаматдегідрогеназна реакція приймає участь не стільки в окисленні глутамата, скільки у синтезі його з б-кетоглутарату. Вона забезпечуює безперервне перетворення вільного аміаку на аміноазот амінокислот. Основний шлях окислення глутамата в мозку полягає у трансамінуванні. У мітохондріях мозку до 90% глутамата трансамінується з утворенням аспартату. Аспартатамінотрансфераза — фермент, який каталізує трансамінування глутамата з оксалоацетатом, є найбільш потужною трансаміназою головного мозку. Виділено два ізоферменти аспартатамінотрасферази, які локалізовані у мітохондріях і цитоплазмі.
Рисунок 4. Утворення й окислення глутамата в головному мозку (1 — глутаматдегідрогеназа; 2 — аспартатамінотрансфераза; 3 — аланінамінотрансфераза; 4 — тирозинамінотрансфераза; 5 — трансаміназа ГАМК)
Мітохондріальний фермент пов’язаний із функціонуванням ЦТК, цитоплазматичний визначає інтенсивність глюконеогенеза. Шлях метаболізму глутамату через трансамінування набагато активніший за дегідрогеназний. У регуляції співвідношення між цими двома шляхами, які конкурують за один і той самий субстрат, важлива роль належить макроергічним сполукам, що сприяють перетворенню НАДФ+ на НАДФН2 і тим самим пригнічують дезамінування глутамату. Трансаміназний шлях потребує використання макроергічних сполук, тому вибір між цими двома реакціями визначається енергетичними властивостями мітохондрій. При нормальному функціонуванні ЦТК дегідрогеназний шлях окислення глутамата пригнічений, а трансаміназний активно протікає. Внаслідок зменшення кількості макроергічних сполук, наприклад, при додаванні до мітохондрій роз'єднувача окислювального фософрилювання 2,4- динітрофенола, пригнічується трансаміназний і значно активується дегідрогеназний шлях окислення глутамату. Взаємоперетворення б-кетоглутарату і глутамату протікає дуже швидко.
У головному мозку був ідентифікований метаболічний шлях такого взаємоперетворення, що отримав назву аспартат-малатного шунта, який служить для транспорту відновлювальних еквівалентів із цитизолю у мітохондрії (рис.5) Потік аспартату через мітохондріальну мембрану пов’язаний з потоком глутамата у зворотному напрямку; також реципрокно пов’язаний потік малату і б-кетоглутарату. Ферменти, що каталізують окремі реакції малат-аспартатного шунта, переважають у тканинах ЦНС.
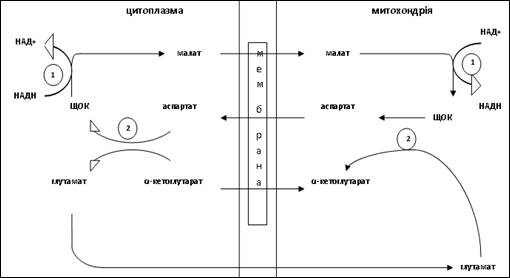
Рисунок 5. Метаболічні компоненти малат-аспартатного шунта (1 -малатдегідрогеназа; 2 — аспартатамінотрансфераза).
Глутамінова кислота виконує дуже важливу функцію в енергетичному забезпеченні головного мозку, яка полягає у підтримці метаболітів ЦТК на достатньо високому рівні, а також забезпеченні мітохондріальних синаптичних процесів відновлювальними еквівалентами. Велике значення має утворення аміаку з глутамата. За участю численних амінотрансфераз аміногрупи різних амінокислот переносяться на глутамінову кислоту, яка трансамінується із оксалоацетатом за участю аспартатамінотрансферази з утворенням аспартата. Утворення аміака з аспартата протікає по різному у мітохондріях і цитоплазмі. У мітохондріях цей процес пов’язаний з амінуванням дезаміноформ НАД (ДНАД) і включає у себе три ферментативні реакції. Поза мітохондріями протікає інший циклічний процес утворення аміака, в якому аспартат реамінує інозинмонофосфат. Для ліквідації аміака у ЦНС, служить глутамінсинтетазна реакція. Глутамінсинтетаза каталізує реакцію:
глутамінсинтетаза.
NH3++АТФ глутамінова кислота глутамін + АДФ + Фн + Н2О.
Mg2+
Цей фермент в мозку тварин знаходиться у більш високій концентрації ніж в інших органах і складає 0,2% від загального білка мозку. У фізіологічних умовах, коли є достатній рівень АТФ, глутамінсинтетазна реакція спрямована у бік зв’язування аміака.
Утворення глутаміна є важливим механізмом детоксикації амонію, до якого мозок дуже чутливий, і накопичення якого шкідливе для ЦНС. Основна частина глутамінсинтетази локалізована у гліальних клітинах й тільки невелика її частина представлена у нервових закінченнях. Дезамінування глутаміну каталізується глутаміназою — ферментом, який локалізований у мітохондріях нейронів. Активність цього ферменту у головному мозку невелика, продукти реакції - глутамінова кислота і амоній гальмують активність фермента. Глутаміназа відіграє важливу роль також в регуляції вмісту глутамата в нервових закінченнях. Глутамінсинтетаза локалізована в основному в гліальних клітинах, а глутаміназа найбільш активна в нейронах. Глутамін є головним попередником глутамата і ГАМК, які виконують нейротрансмітерну функцію. Тому виникла концепція про існування глутамінового цикла. Глутамат поглинається гліальними клітинами, перетворюється в глутамін у синтетазній реакції, останній надходить в нейрони та утворює глутамінову кислоту. Таким чином, глутамін служить гліально-нейрональним транспортером глутамата.
Рисунок 6. Утворення аміаку у циклі реамінування Д-НАД (1 — НАД-сукцинатсинтетаза; 2 — НАД-сукцинатліаза; 3 — дезаміназа НАД)
Важливою функцією глутамата є участь в біосинтезі білків і біологічно активних пептидів. Глутамат і глутамін складають разом від 8 до 10% загальних амінокислотних залишків у гідролізаті білків мозку. Глутамат є складовою частиною ряду малих і середніх регуляторних пептидів мозку: глутатіону і ряда б-глутамільних дипептидів. Деякі нейропептиди містять циклічні похідні глутамата — піроглутамат, люліберин, тироліберин, нейротензин, бомбензин та ін. Основним джерелом нейротрансмітерного глутаматного фонду є глутамін, який синтезується переважно в астроцитах, де локалізована глутамінсинтетаза. Глутамін легко транспортується через мембрану астроцитів і за допомогою активних носіїв досягає нервових закінчень.
Рисунок 7. Утворення аміаку у циклі реамінування ІМФ (1 — аденілсукцинатсинтетаза; 2 — аденілсукцинатліаза; 3 — аденілатдезаміназа)