Фіксація атмосферного азоту вільноживучими і симбіотичними азот фіксаторами

Клубочкові бактерії дуже чутливі до низького значення рН ґрунтового розчину. Так, конюшина може добре рости при рН 4, якщо його постачати мінеральним азотом, однак його клубочкові бактерії нездатні до азотфіксації. Клубочкові бактерії люпину, конюшини і квасолі характеризуються найбільшою кислотністю, а в люцерни низька границя рН лежить у інтервалі 4,8−5,2. Внесок концентрації нітратів і амонію… Читати ще >
Фіксація атмосферного азоту вільноживучими і симбіотичними азот фіксаторами (реферат, курсова, диплом, контрольна)
Фактори які впливають на фіксацію молекулярного азоту.
Клубочкові бактерії володіють здатністю асимілювати значну кількість атмосферного азоту і постачати бобовим рослинам. Добрі посіви конюшини і люцерни можуть зв’язати у рік до 400кг/га азоту. В даний час відомо приблизно 200 видів бульбучкових бактерій, які здатні до азотфіксації і представляють інтерес для землеробства.
Клубочкові бактерії дуже чутливі до низького значення рН ґрунтового розчину. Так, конюшина може добре рости при рН 4, якщо його постачати мінеральним азотом, однак його клубочкові бактерії нездатні до азотфіксації. Клубочкові бактерії люпину, конюшини і квасолі характеризуються найбільшою кислотністю, а в люцерни низька границя рН лежить у інтервалі 4,8−5,2. Внесок концентрації нітратів і амонію в поживному середовищі подавляють утворення на коренях бульбочок і знижують їх активність. Мобілізація азоту збільшується при підвищенні температури до 22оС, максимальна при 26−27оС. Найбільш активно проходить цей процес у верхніх горизонтах ґрунту, володіючи високою біологічною активністю. Так, по даних П. М. Смирнова на дерново-підзолистих ґрунтах з непорушеною структурою в товщі шару 40−85см. закріпилося в органічні форми лише 1−4% внесеного азоту. В слої промивного водного режиму викликати рух нітратів в глибоких горизонтах, розміри мобілізації мінерального азоту знижуються, складає у розрахунку на 165 — сантиметрову товщу за 4,5 місяці приблизно 17%. У виді нітратів у профілі ґрунту виявлено 40% внесеного азоту, засвоєння його рослинами склало 34%. Втрати цього елемента живлення в умовах вимивання нітратів не перевищує 9%.
На фіксацію азоту впливає такий процес, як обробка насіння бобових культур перед посівом на основі бульбочкових бактерій виготовляють препарати нітрагін, нітразон, фосфоробактерії. Існують різні типи нітрагіну в залежності від способу його приготування: суміш бактерій із сухими наповнювачами для обробки насіння і водна суспензія бактерій для обприскування; препарат для інокуляції насіння методом вакуумної інфільтрації. Мікробіологічна промисловість випускала дві форми нітрагіну (сухий нітрагін) і ризоторфін (торф'яний нітрагін). Технологічна схема виробництва сухого нітрагіну включає наступні етапи: одержання посівного матеріалу > ферментація > зміщування рідкої культури з захисним середовищем > висушування розпилюючим способом.
Гектарна порція ризоторфіну — 200 г. Передпосівна обробка насіння здійснюється шляхом розведення препарату у воді і перемішування його з насінням в день посіву. Висів обробленого ризоторфіном насіння необхідно здійснювати у вологий ґрунт, удобрений фосфором, калієм і мікрокислотами, особливо молібден. На ґрунтах слабокультурних, бідних на азот, застосування ризоторфіну варто поєднати з внесенням невеликих доз мінерального азоту не більше 45кг/га. Інокуляція бобових бульбочковими бактеріями особливо ефективна на площах, де раніше не вирощувалася дана бобова культура і відповідні специфічні бактерії в ґрунті відсутні.
На фіксацію азоту впливають макроі мікроелементи. Наприклад, мікроелемент сірка відіграє важливу роль в окисленні і відновлених процесах, активізації ферментів, синтез білків. Сірка бере участь в асиміляції нітратів рослиною стимулює її нагромадження. При нестачі сірки під час живлення рослин затримується синтез білків і нагромадження азоту у небілкові форми, нітратів. У бобових культур знижується життєдіяльність бульбочкових бактерій і синтез хлорофілу. Під впливом сірки збільшується не тільки урожай, а й покращується якість продукції і вміст білка може збільшуватися 2%. Загальний вміст сірки коливається від 20−35мг/м в ґрунті.
Молібден потрібний для живлення бобових культур і фіксації молекулярного азоту із повітря. Молібден є складовою частиною ферменту нітрат редуктоз, які беруть участь у відновленні нітратів у частинках кореня до аміаку.
Важливий фактор який впливає є світло. Має прямий зв’язок із живлення, зміна дня і ночі, зміна вологості, температури. Інтенсивність освітлення це виробилося у рослин і тому вони постійно реагують і як наслідок нормального розвитку.
Також впливає на фіксацію азоту механічний склад ґрунту, фізичні властивості, кількість гумусу, глибина орного шару, вологість ґрунту, пористість, щільність, аерація.
Симбіотичні азотфіксатори
Бульбочки утворюються тільки у представників родини бобових (Fabaceae). У різних рослин бульбочки розрізняються тільки за формою і величиною. Утворюються вони після проникнення в кореневу систему бульбочкових бактерій.
Старіючи, бульбочкові бактерії втрачають джгутики, перестають бути рухливими і набувають вигляду оперезаних паличок, оскільки з віком бактеріальна клітина наповнюється жировими включеннями, які не забарвлюються. Зі старінням у бульбочках культури Rhizobium часто виникають потовщені, розгалужені, сферичні та іншої форми утворення, які значно більші за звичайні клітини. Ці поліморфні утворення дістали назву бактероїдів (рис. 1). Вони нерухомі і не здатні до розмноження. Проте найбільш енергійно відбувається засвоєння азоту в бульбочках саме тоді, коли бульбочкові бактерії перетворюються на бактероїди.
Встановлено, що бульбочкові бактерії можуть заражати лише певну групу бобових рослин. Вибіркова здатність цих бактерій відносно рослин дістала назву специфічності. Ця властивість стала головною ознакою для розробки систематики бульбочкових бактерій.
За Л. М. Доросинським, рід Rhizobiumподіляють на такі групи: R. leguminosarum— бактерії гороху, вики, кормових бобів, чини; R. phaseoli— квасолі; R. japonicum— сої; R. vigna— вігни, арахісу, машу; R. сісеr — нугу; R. lupini— люпину, серадели; R. trifolii— еспарцету; R. lotus— лядвенцю; R. robinii— акації.
В окремих випадках спостерігається не тільки видова, а й сортова специфічність бульбочкових бактерій. Крім специфічності, цим бактеріям властива вірулентність — здатність проникати в тканину кореня, розмножуватися там і спричиняти утворення бульбочок. За певних умов ці бактерії можуть знижувати або зовсім втрачати активність.
Істотною властивістю бульбочкових бактерій є також їхня активність, тобто здатність у симбіозі з рослинами асимілювати молекулярний азот. У ґрунті трапляються штами активних і неактивних бульбочкових бактерій. Зараження бобових рослин активною расою бактерій веде до утворення великої кількості бульбочок на головному корені та зумовлює енергійний процес фіксації атмосферного азоту. Неактивні раси цих бактерій спричинюють утворення бульбочок, але азот не фіксується.
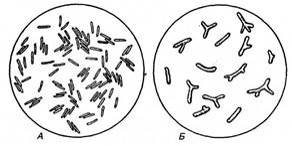
Рис. 3.1. Бульбочкові бактерії з роду Rhizobium:
А — клітини, виділені з бульбочок конюшини; X15 000−25 000 (за Зільгером); Б — бактероїди з бульбочок конюшини Бульбочки, які утворюються активними расами бактерій, мають рожеве забарвлення. Пігмент, що надає їм такого забарвлення, за хімічним складом близький до гемоглобіну крові та називається леггемоглобіном (фітоглобін). Вважають, що цей пігмент сприяє процесу засвоєння азоту, підтримуючи окислювально-відновний потенціал на певному рівні. Бульбочки, які утворюють неактивні раси бактерій, мають зеленкуватий колір.
Механізм проникнення бактерій у кореневий волосок досі ще недостатньо вивчено. Встановлено, що на поверхні клітинної оболонки бобових рослин є особливий вид білка, до якого вибірково «приклеюються» специфічні бульбочкові бактерії завдяки наявності в їхній, оболонці специфічного ліпополісахариду. Припускають, що саме у відповідь на появу цього ліпополісахариду на оболонці кореневого волоска з’являється білок лектин, який зв’язує полісахариди оболонки ризобіїв. Це своєрідний механізм «впізнавання» перед виникненням бобово-ризобіального симбіозу.
Після проникнення бактерій у клітину кореневого волоска вони починають посилено розмножуватись і утворюють суцільний тяж, так звану інфекційну нитку (слизовий тяж), в якій скупчується величезна кількість бульбочкових бактерій. Останні можуть розмножуватися тільки в тетраплоїдних клітинах рослин.
Як в інфікованих бактеріями, так і в сусідніх клітинах починається активний поділ, і утворюються бульбочки. В однорічних рослин бульбочки є тимчасовим утворенням. У багаторічних вони можуть функціонувати протягом кількох років. Контакт бактерії з рослиною встановлюється через судинні пучки, які обплітають тканину бульбочки. Провідними шляхами до бульбочок надходять вуглеводи та інші поживні речовини, де вони використовуються бактеріями, а рослина одержує із бульбочок зв’язані сполуки азоту. Доведено, що найінтенсивніша віддача зв’язаного азоту із тканин бульбочки відбувається тоді, коли бактероїди перебувають у життєдіяльному стані. Транспорт азотовмісних речовин із бульбочок у рослину здійснюється у формі амінокислот. Незначна кількість засвоєного рослиною азоту виділяється внаслідок екзоосмосу коренями в ґрунті переважно у вигляді аспарагінової кислоти.
Після відмирання і розкладу бульбочок бактерії потрапляють у ґрунт і живуть там як сапрофіти, поки знову не проникнуть у корені рослин. Питання про те, що лежить в основі взаємовідносин бактерій і бобових рослин (паразитизм чи співжиття), ще повністю не з’ясовано. Важливим є той факт, що бульбочкові бактерії, проникаючи в рослину, стимулюють розвиток у неї імунітету, який перешкоджає подальшому зараженню коренів.
Отже, вклад бобових рослин у забезпечення ґрунтів доступними формами азоту досить відчутний.
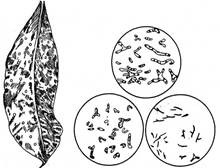
Рис. 3.2. Бульбочки на листках Pavetta.
Утворення, які нагадують бульбочки, знайдено у багатьох рослин, що не належать до родини бобових. Серед покритонасінних бульбочки виявлено на коренях представників порядків Cycadales, Ginkgoales і Coniferales. У дводольних бульбочки найчастіше бувають у представників родин Согіагіасеае, Betulaceae, Casuarinaceae тощо. В окремих випадках їх знаходили на коренях рослин із родин Brassicaceae, Rosaceae, Scrophulariaceae тощо, з однодольних рослин бульбочки описано лише у представників родини Роасеае.
Нещодавно з’явилися цікаві дані про те, що бактерії роду Azospirillum, які асоціюються з коренями злаків, мають здатність інтенсивно фіксувати азот. Вони поширені в тропічних і помірних кліматичних зонах. Природу збудників, які зумовлюють утворення бульбочок у цих рослин, повністю ще не вивчено. Так, у бульбочках вільхи знайдено актиноміцети.
Досліди з використанням міченого азоту показали, що у більшості рослин із родів вільхи (AlnusL.), обліпихи (HippophaeL.), лоху (ElaeagnusL.) мічений азот зв’язується тільки тоді, коли є бульбочки. Ці дані свідчать про те, що зв’язування молекулярного азоту небобовими рослинами також відбувається в симбіозі з мікроорганізмами.
Бульбочки можуть утворюватися не тільки на коренях рослин, а й на інших органах. їх знайдено на листках майже 400 різних видів рослин, наприклад деяких дводольних тропічних, зокрема у павети (PavettaindicaL.) із родини маренових (Rubiaceae). Найдокладніше досліджено такі бульбочки у рослин павети і психотрії (рис. 4).
Вільноживучі азот фіксатори
Крім бульбочкових бактерій, у ґрунті є багато інших видів мікроорганізмів, які можуть засвоювати молекулярний азот атмосфери. В 1893 р. С. М. Виноградський вперше виділив і вивчив вільноживучий азотфіксатор — анаеробну спороносну бактерію веретеноподібної форми, яку було названо на честь Л. Пастера — Clostridiumpasteurianum.
Описано багато азотфіксаторів з роду Clostridium (C.buturicum, C. acetobutylicum, C. pectinovorum, C. felsineum, Cl. beijerinckii та ін.). Ці бактерії можуть використовувати різні джерела азоту: солі амонію і азотної кислоти, а також багато різних органічних азотовмісних сполук. Із вуглецевих сполук вони використовують моноцукри, дицукри, поліцукри, органічні кислоти тощо.
Енергійним фіксатором азоту серед цієї групи бактерій є Clostridiumpasteurianum. Він може зв’язувати до 10—12 мг азоту на 1 г збродженого цукру.
Іншим дуже поширеним вільноживучим азотфіксатором є аеробна, овальної форми, бактерія — Azotobacterchroococcum, відкрита у 1901 p. M. Бейєрінком. Розмір клітин азотобактера коливається в межах 2—3×4—6 мкм. Він розмножується простим поділом з утворенням поперечної перегородки. Молоді клітини азотобактера рухливі, перетрихи з віком втрачають рухливість, набувають майже коковидної форми і покриваються товстим шаром слизу (капсулою). Іноді клітини азотобактера можуть бути вкритими товстою оболонкою і перетворюватися на цисти.
Серед представників азотобактера найґрунтовніше вивчено A. chroococcum, A. vinelandii, A. agilis, A. beijerinckii (рис. 3). Ці види різняться за формою і розмірами клітин, пігментацією колоній.
На відміну від клостридія азотобактер інтенсивніше зв’язує молекулярний азот. Активні культури азотобактера зв’язують 15—20 мг азоту на 1 г використаного цукру або іншої органічної речовини. Він не засвоює клітковини. Однак при наявності її в ґрунті розмножується більш інтенсивно, оскільки між целюлозорокладачами і азотобактером існує явище метабіозу. Азотобактер потребує нейтрального середовища. В кислих ґрунтах він не розвивається.
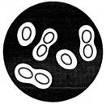
Рис. 3.3. Азотобактер (Azotobacterchroococcum)
Близькі до азотобактера вільноживучі азотфіксатори з роду Beijerinckia. На відміну від азотобактера, вони можуть рости навіть при рН = 3. Ці бактерії мають різну форму, бувають рухливими і нерухливими; за енергією фіксації азоту вони близькі до азотобактера.
Серед вільноживучих азотфіксуючих бактерій слід згадати про види роду Derxia і Azotomonasfluorescens.
Пошуки вільноживучих азотфіксаторів дали змогу виявити їх і в інших родинах — Spirillaceae, Rhizobiaceae, Achromobacteriaceae, Enterobacteriaceae, Bacillaceae та ін. Здатність до фіксації молекулярного азоту мають анаеробні пурпурні та зелені фототрофні бактерії, що живуть у заболочених озерах і мулі, деякі мікобактерії, спірохети, проактиноміцети і навіть окремі види грибів, у тому числі дріжджі.Використання методу мічених атомів дозволило підтвердити припущення, висловлене ще в 1889 p., про те, що одна з найбільших груп мікроорганізмів, які населяють ґрунти, — мікроскопічні водорості - можуть засвоювати атмосферний азот. Серед них близько 40 видів синьозелених водоростей. До найпоширеніших належать Anabaena, Amorphonostoc, Aulosira, Calothrix, Nostoc, Scytonema тощо.
В Японії використання штучного зараження ґрунтів синьозеленими водоростями Tolypothrixtennuisвиявилось ефективним при вирощуванні рису. Подібних результатів досягли також в Індії від застосування на рисових полях синьозелених водоростей — Aulosirafertilisima.
Досліди, проведені в Інституті мікробіології Російської АН, показали, що синьозелені водорості (ціанобактерії) під час інтенсивного розвитку можуть нагромаджувати за вегетаційний період до 50—70 кг азоту на 1 га.
Останніми роками вчені різних країн приділяють велику увагу вивченню процесу фіксації азоту мікроорганізмами, які містяться на корінні і в прикореневій зоні небобових рослин. Ці мікроби дістали назву ризосферних, а процес зв’язування ними молекулярного азоту називається асоціативною азотфіксацією.
Азотфіксуюча активність виявлена у представників багатьох родів ризосферних бактерій: Agrobacterium, Achromobacter, Aquaspi-rillum, Azospirillum, Arthrobacter, Bacillus, Flavobacterium, Enterobac-ter, Erwina, Flavobacterium, Klebsiella, Mycobacterium, Pseudomonas, Rhodospirillumта ін.